How x-ray diffraction with synchrotron radiation got started
The need to record low angle scattering x-ray fibre diagrams from muscle with milli-second time resolution drove the use of synchrotron radiation as an x-ray light source. The first smudgy diffraction patterns were obtained from a slice of insect flight muscle. Out of this grew the EMBL Outstation at DESY.
Synchrotron radiation has proved to be of inestimable importance for extending the scope of x-ray diffraction methodology for protein crystallography. The laser-like optics and tremendous brilliance allow x-ray diffraction data to be collected from ever smaller crystals of ever larger complexes and organelles. In addition, the ability to select wavelengths makes it possible to exploit anomalous scattering for phase determination (MAD). Moreover, the continuous spectrum makes it possible, in favourable cases, to obtain data in fractions of a second. Within the next decade genome sequencing projects will provide us with a complete menu of all DNA, RNA, and protein molecules in a number of organisms and will thereby give us the inventory of life. To make full use of all these data we will need to understand the macromolecular interactions which control cell signalling, cell locomotion and gene activation. To do this we will need a detailed description of the structure of many of the relevant macromolecules. At present, only x-ray protein crystallography yields a molecular anatomy of adequate resolution and precision for this task. Thus there would seem to be an almost limitless need for x-ray crystallography groups working on proteins if full use is to be made of the flood of genetic information arising from the sequencing projects.
However, this is not how x-ray diffraction with synchrotron light got started, it started with muscle.
Muscle - the Problem
Muscle is an isothermal engine which works by hydrolysing ATP (adenosine triphosphate) with an efficiency near 50%. When muscle contracts, two sets of protein filaments, the actin filaments and the myosin filaments glide over each other. The gliding is driven by the cyclical interactions of the myosin "cross-bridges" with actin so as to "row" one set of filaments past the other: a cross-bridge binds to actin in a initial position and "swings" into a final position. This movement is driven by the binding to actin which enables release of the products of ATP hydrolysis (ADP and phosphate). At the end of the stroke ATP rebinds to the myosin cross bridge causing rapid release from actin. Subequently ATP is hydrolyzed to ADP and phosphate and the cycle repeats (Fig 1) (Lymn & Taylor, 1971). One major aim of muscle research has been to demonstrate and understand what actually happens when a cross bridge "swings".
The cross bridges in muscle fibres repeat along the fibre axis with a repeat distance of ca. 14.5nm. Thus they give rise to an x-ray diffraction pattern with a series of strong meridional reflexions. Alterations in shape of the cross bridges lead to changes in the intensities of these reflexions. The sartorius muscle from frog can be dissected out intact and made to contract by electrical stimulation. X-ray diffraction patterns can then be recorded from an actively contracting muscle. Pioneering work with conventional sources was carried out in the 1960's by H.E. Huxley (Fig 2) (Huxley & Brown, 1967). However, the scattering is weak and muscles quickly become fatigued. We in Heidelberg hoped that insect flight muscle might provide an alternative system to frog muscle for studying the cross bridges by x-ray diffraction. Insect flight muscle changes its structure on adding ATP which leads to dramatic changes in the low angle meridional reflexions (Reedy et al ., 1965). Moreover, it is highly crystalline. However, we met all the intensity problems encountered by H. E. Huxley for frog muscle but in a more acute form since the specimens are much smaller. To get further we needed much stronger x-ray sources.
First diffraction experiments are carried out at DESY
In 1969 Rosenbaum started his doctorate in Heidelberg on developing x-ray sources for diffraction studies of muscle Together with H.E. Huxley in Cambridge a rotating anode x-ray tube of high brilliance was developed (which became the GX18 of Elliott Bros). However, one quickly runs into a performance barrier set by the strength of materials (Rosenbaum, 1979). Frustration with rotating anode tubes drove us to re-evaluate the performance of synchrotrons as x-ray sources. Initial estimates of the strength of synchrotron radiation K.C. Holmes had made in the '60s indicated that synchrotrons at 6.0 GeV would not be much better than existing rotating anode tubes. In the mean time the energy of the machine had risen from 6.0GeV to 7.2GeV and the beam current was often in excess of 10mA. Now things looked more promising. Moreover, G. Rosenbaum had done his diploma work at DESY in the F41 VUV group so that he could provide important know-how about the physical set up at DESY. Therefore, with the ecouragement of Dr Heansel and the F41 group we set about conducting trials in the VUV bunker at DESY.
Working together with Jean Witz, who was an authority on x-ray optical systems, we introduced a focusing x-ray quartz monochromator into the synchrotron beam. A vacuum chamber was constructed to house a bent quartz monochromator and slit assemblies with a berillium exit window (Fig. 3). The quartz monochromator could be moved across the beam by remote control. For all other adjustments the main beam shutter had to be closed: reopening the beam shutter required retreating behind a massive sheilding wall, setting up the interlocks, and a telephone call to the main control room. This process made adjustment rather tedious. However, estimates of the monochromatic beam intensity were very encouraging. Therefore we set up a primitive x-ray diffraction camera on the monochromatic beam and the first rather smudged x-ray diffraction photograph using synchrotron radiation (from a strip of insect flight muscle kindly supplied by Dr. H-G. Mannherz) was obtained in August 1970 (Rosenbaum et al ., 1971).
Bunker-2
On the basis of these initial experiments the directors of DESY (in particular Martin Teucher, who was responsible for buildings) encouraged us to set up a bunker for x-ray diffraction experiments on biological samples. The original (and final) plan was to set up an x-ray lab on the storage ring DORIS which would have at least 100 times more intensity than DESY. However, we were persuaded by Prof. Jenschke, the founding director of DESY that since DORIS would not be available as a synchrotron light source for 2-3 years it would be advantageous to build an x-ray laboratory on the DESY synchrotron in order to gain experience. However, we would need to act fast! The window of opportunity was the major shut-down in 1971 engendered by the construction of the connecting tunnels from DESY to the new strorage ring DORIS. After this date the massive earth movements required to create a new bunker would no longer be possible. Thus an x-ray laboratory was built onto DESY during the shut down of 1971 and became known as "Bunker-2". At the same time the tunnel for the laboratory-to-be on DORIS were also built (to be completed in 1975). Above Bunker 2 two offices and a room for biochemistry were added.
EMBL


Fig 4: In 1975 EMBL and DESY entered into a formal agreement to set up an EMBL outstation at DESY in Hamburg. The photograph shows H. Schopper Chairman of the Directors of DESY, J.C. Kendrew Director General of EMBL and W. Berghaus the Administrative Director of DESY at the signing ceremony.
The history of this bunker became entwined with the history of the European Molecular Biology Laboratory, EMBL. It was clear that a sychrotron radiation x-ray diffraction facility would become a central facility for European research, particularly in molecular biology where the advantages of good x-ray optics and intensity were appreciated much earlier by the structural biologists than in the general crystallographic community, who only became interested a decade later. What better vehicle for such a facility could there be than EMBL. In 1969 advised by H.E. Huxley, we made a proposal for such a facility to Sir John Kendrew, the head of the EMBL "Project" (EMBL did not exist legally for another 5 years). John Kendrew greeted our initiative with enthusiasm. The new EMBL laboratory was dedicated to technological developments for molecular biology. High level contacts ensued at which it was agreed that the developments in Hamburg should become part of an outstation of EMBL at DESY. Thus a little later Bunker 2 became the provisional headquarters of the EMBL outstation. The initial financing of this project is a tribute what can be achieved through good will. Salaries were initially paid by the Deutche Forschungsgemeinschaft and then by EMBO (the private sister organisation to EMBL). Building costs were covered by the Bundesministerium für Wissenschaft und Forschung (via DESY) - perhaps hoping this would help ensure that EMBL was established in Germany - and the Max Planck Institute for Medical Research in Heidelberg carried the equipment costs. With the ratification of the EMBL agreement in 1974 the whole project was taken over by EMBL. An official agreement between DESY and EMBL setting up the Outstation was signed in 1975.
The first beam line
In the mean time work went on. Inside Bunker 2 a massive neutron-proof concrete wall separated the operators from the beam. Therefore, all adjustments had to be made by remote control. In collaboration with John Barrington Leigh, Gerd Rosenbaum set about building a fully remotely controlled optical bench (Barrington Leigh & Rosenbaum, 1974; Barrington Leigh & Rosenbaum, 1976) (Fig. 5). A Guinier monochromator was used to focus the fan of radiation from the synchrotron in the horizontal plane and 2 x 20 cm. adjustable bent mirrors were used to focus the much smaller divergence in the vertical plane. The mirrors (fused quartz) were nearest to the synchrotron and were housed in a helium-filled box separated from the machine vacuum by a beryllium window. Beams were accommodated in vacuum tubes fitted with mylar windows. It proved difficult to obtain mirrors polished to the necessary flatness: optical mirror manufacturers had no way of monitoring the micro-flatness necessary for x-ray mirrors. Here we were considerably helped by the pioneering work of Franks (Franks & Breakwell, 1974). Movements were controlled by about 100 small DC motors with potentiometers as position sensors. DC motors were chosen rather than stepping motors because they are light: the whole apparatus was built on a mini budget and the apparatus could not become massive. A SIT-vidicon camera was used to observe in line the image of the direct beam formed on a cesium iodide crystal. Two other steerable TV cameras fitted with zinc sulfide screens were mounted on a parallel optical bench for visual observation. Zinc sulfide screens could be inserted into the beam path by remote control for monitoring the beam e.g. before and after the slits. The monochromator (quartz) was cut at 7° to the surface so as to approximate to the Guinier-condition for the given source distance and the desired focal distance, i.e. the desired demagnification. The deviation from the exact Guinier-geometry resulted in a wavelength inhomogeneity across the converging beam. However, this effect was small for the apertures being used and was not important in small-angle diffraction. The angle of the latter part of the optical bank to the direct beam was fixed for _=1.5Å. Since the optical elements were about 40m from the tangent-point of the synchrotron and focused within 2-3m a demagnification of H 15 was achieved.
The electron beam of DESY was relatively compact so that a focused beam of dimensions 200 x 250 m could be obtained. With a flux of H 10 9 photons/s and excellent optical properties this was a very good beam for low angle scattering. The flux density was two orders of magnitude better than could be achieved with the best rotating anode tubes. Images were registered on film or on one-dimensional single-wire (Gabriel & Dupont, 1972) position sensitive detectors. The beam line was in operation in 1972 and for a couple of years remained a unique facility.
Diffraction from insect flight muscle
Using this beam line, the Heidelberg group (in collaboration with Richard Tregear from Oxford) studied the diffraction from insect flight muscle. The excellent collimation led to detailed fibre diffraction pictures which yielded new structural information (Holmes et al ., 1980). Time resolved experiments were set up with oscillating insect flight muscle. The muscles were attached to a vibrator and oscillated at 5 Herz at which frequency they generate considerable work if provided with ATP. At low amplitudes of oscillation it was expected that the cross bridges might be partially synchronized so that one should be able to record diffraction patterns from various parts of the cross-bridge cycle. The diffraction was recorded, a layer line at a time, on a position sensitive detector and the output switched into one of 32 bins in synchrony with the oscillation. Data with usable statistics could be obtained from the equator in about 15mins. However, on account of the available intensity, the measurements remained confined to the strong equatorial reflexions (Barrington Leigh & Rosenbaum, 1976). Unfortunately, these reflexions alter little between resting and contracting insect flight muscle and, therefore, are not very useful for monitoring the cross-bridge swing. We were not observing the cross bridges in flagranti . The insect flight muscle experiments needed a storage ring!
Since the intensity was not adequate to allow a time resolved study of the meridional reflexions (which do alter with cross bridge orientation) attempts were made to "freeze" the cross-bridges in alternative conformations by the use of non-hydrolysable analogs of ATP (Goody et al ., 1976). Quite large changes in the diffraction pattern were induced by certain analogs. However, a large part of the changes resulted from alterations in the pattern of binding of the cross bridges to actin, rather than in an underlying change in the cross-bridge orientation (Goody et al ., 1975).
The group shared experiences with Hugh Huxley and Uli Arndt in Cambridge who were setting up a similar beam line at NINA for experiments on frog muscle.
Protein crystallography
The initial success at DESY sparked world wide interest. In June 1972 there was a historical meeting in Brookhaven at which most of the subsequent applications of x-ray synchrotron radiation were discussed (see -BNLReport 1973). The most important application for biology later proved to be protein crystallography. Early tests of protein diffraction on the DESY source (Harmsen et al ., 1976) showed improvements compared with conventional sources but the gains were limited. The flux was about ten times better than with a conventional source. At this stage one had failed to appreciate that the parallel collimation of the beam was giving unusually good signal/noise. This was the property of synchrotron radiation which ultimately made it the source of choice for all kinds of protein crystal data collection. About the same time studies on the Stanford storage ring SPEAR (Phillips et al ., 1976) showed gains for crystal diffraction even with a non-focusing monochromator which showed clearly that storage ring sources were to be of considerable importance in protein crystallography. These authors made use of the ability to "tune" the wavelength across an adsorption edge to demonstrate the potentialities of synchrotron radiation in exploiting the effects of anomalous dispersion.
Bunker 4
The experimental facility at DORIS was housed in a small experimental hall rather than a bunker. Nevertheless, it was known as "Bunker-4". Duly equipped with offices, seminar room, workshops, and a biochemistry laboratory this building became the home of the EMBL Outstation in 1975. DORIS is a colliding beam facility with electron and positrons circulating in opposite directions. The beams into the EMBL bunker were from the positron ring. The first beam-line set up (X11, designed by G. Rosenbaum and A. Harmsen, later taken over by H. Bartunik) was a mirror-monochromator combination with 8x20cm mirrors and a bent germanium monochromator (Rosenbaum, 1979; Rosenbaum & Holmes, 1980). The bench carrying the specimen and detector could be rotated around the monochromator as pivot so as to vary the wavelength. Each of the mirrors could be individually bent. The electron beam in DORIS was considerably larger in cross-section than that of DESY so that fine-focused beams such as were obtained on the DESY synchrotronwere not attainable. In fact it turned out that although each of the mirrors was designed to be individually bent it was not really worth bending the mirror segments at all, aligning them appropriately without bending produced as fine a beam as one could get. A second optical system X13 (Bordas et al ., 1980) similar in design to X11 was soon added. The two shared a common mirror box and mirror design. These beam lines were the workhorses of the DORIS facility for a number of years. The EMBL Outstation in Bunker-4 expanded steadily and became one of the most widely used biological facilities in the world.
Time resolved studies on frog muscle
HE Huxley continued with frog muscle and transferred his work to the EMBL Outstation when this became operative (Huxley et al ., 1980). The key experiments were carried out a little later (Huxley et al ., 1981) by a team which included a number of people who have subsequently played an important role in the development of synchrotron radiation as an x-ray light source. These experiments finally showed the anticipated changes in intensity of the meridional reflexions: if a contracting muscle is released the intensity of the 14.35nm meridional reflexion drops within a few ms to a fraction of its initial value. If the muscle is extended quickly, the intensity is recovered. If one waits at the new length the intensity recovers. These experiments have recently been repeated with very high time resolution using sophisticated mechanics and the excellent two dimensional detectors at Daresbury (Irving et al ., 1992). These observations are fully consistent with the swinging cross bridge hypothesis and for many years represented the most important time resolved experiments supporting the swinging cross-bridge hypothesis.
Unfortunately, the resolution of these changes is actually too low to show the detailed molecular basis of the cross-bridge swing. Over the years it became likely that the swinging cross bridge was actually a swinging lever arm (Fig 6) (Cooke, 1986). Protein crystallography, which in turn needed synchrotron light, finally yielded insight into this mechanism.
Atomic structures give insight
The structure of the actin monomer and of the actin filament have been solved by protein crystallography (Kabsch et al ., 1990) and by x-ray fiber diffraction (Holmes et al ., 1990) . The crystal structure of the myosin subfragment 1 (Rayment et al ., 1993b) showed the myosin cross-bridge to have an extended C-terminal neck which looked like the anticipated lever arm and, moreover, a lever arm which was in the correct orientation and position to function as a lever arm (Rayment et al ., 1993a). In the last year a number of independent experiments provide results which are in excellent accord with the idea that the C-terminal tail functions as a lever arm and indeed provide evidence that it can move (see review (Holmes, 1997)). Furthermore, new crystal structures (Fisher et al ., 1995; Smith & Rayment, 1996a) with analogs of ATP bound appear to show an alternative orientation of the lever of the anticipated kind.
The crystallographic studies cited show two distinct structural states for the myosin cross-bridge: the "open" or "end" conformation which is characterized by the absence of nucleotide (rigor); and the "closed" or "beginning" state, which is favored by binding ATP or the products complex (ADP.Pi) (Fig 7). Myosin transports actin by switching between these two states. "Open" and "closed" refer to the status of the ATP binding site. This in turn is coupled to the rotation of the C-terminal lever arm. In the "closed" form the lever arm is at the beginning of the power stroke whereas in the "open" form it is at the end of the power stoke. The preference for "open" or "closed" is also controlled by binding to actin. It is likely that the closed state binds only weakly to actin. On this basis the structural states can be correlated with the Lymn-Taylor cycle.
Starting from an actin-myosin complex at the end of the power stroke, the binding of ATP brings about rapid closure of the ATP binding site and concomitant release from actin. The closed state hydrolyses ATP to ADP. Pi without attaching to actin. Thereafter, the rebinding of myosin in the closed or "beginning" conformation of the products complex to actin opens the site to facilitate release of the__-phosphate. Release of phosphate in turn induces an isomerisation to the open "end" conformation since it is the presence of the__-phosphate which stabilizes the closed form. The isomerisation results in large changes of angle of the "lever arm" (at the distal part of the myosin head). Since the S1 is strongly attached to actin at this stage this results in a 12nm transport of actin past myosin.
It would not have been possible without synchrotron light
Thus we see that one of the most important puzzles of biology, the basis of animal movement, which originated as a research project with the Alexandrian school in the third century BC has yielded many of its secrets to a structural and physico-chemical analysis. It is noteworthy that this could not have happened without synchrotron radiation sources. Moreover, this project opened up one of the most important uses of synchrotron radiation yet discovered, namely its use as an x-ray diffraction a source for protein structure determination.
Figures
The Lymn-Taylor cycle
![Fig 1: The Lymn-Taylor cycle (Lymn & Taylor, 1971) . A diagrammatic representation of the crossbridge cycle: the myosin cross bridge is bound to actin in rigor 45°- position - "down" (1) ATP binds which leads to very fast dissociation from actin (2)]. The hydrolysis of ATP to ADP and a phosphate ion leads to a return of the myosin cross bridge to the 90° "up" position(3) whereupon it rebinds to actin (4). This leads to release of the products and return to [1]. In the last step actin is "rowed" past myosin.](/26969/original-1309858144.jpg?t=eyJ3aWR0aCI6MzQxLCJmaWxlX2V4dGVuc2lvbiI6ImpwZyIsIm9ial9pZCI6MjY5Njl9--ba4db6ad2422db0d4792a4b61d9074eb23f05d14 "The myosin cross bridge is bound to actin in rigor 45°- position - down [1]. ATP binds which leads to very fast dissociation from actin [2]. The hydrolysis of ATP to ADP and Pi leads to a return of the myosin cross bridge to the 90° up position whereupon it rebinds to actin [3]. This leads to release of the products and return to [1]. In the last step actin is rowed past myosin.")
![Fig 1: The Lymn-Taylor cycle (Lymn & Taylor, 1971) . A diagrammatic representation of the crossbridge cycle: the myosin cross bridge is bound to actin in rigor 45°- position - "down" (1) ATP binds which leads to very fast dissociation from actin (2)]. The hydrolysis of ATP to ADP and a phosphate ion leads to a return of the myosin cross bridge to the 90° "up" position(3) whereupon it rebinds to actin (4). This leads to release of the products and return to [1]. In the last step actin is "rowed" past myosin.](/26969/original-1309858144.jpg?t=eyJ3aWR0aCI6MjQ2LCJvYmpfaWQiOjI2OTY5fQ%3D%3D--5cd200d49d29f3fe22d2154e929a900db9af2eca)
© Kenneth C. Holmes
Fig 1: The Lymn-Taylor cycle (Lymn & Taylor, 1971) . A diagrammatic representation of the crossbridge cycle: the myosin cross bridge is bound to actin in rigor 45°- position - "down" (1) ATP binds which leads to very fast dissociation from actin (2)]. The hydrolysis of ATP to ADP and a phosphate ion leads to a return of the myosin cross bridge to the 90° "up" position(3) whereupon it rebinds to actin (4). This leads to release of the products and return to [1]. In the last step actin is "rowed" past myosin.
© Kenneth C. Holmes
The Lymn-Taylor cycle (Lymn & Taylor, 1971). A diagrammatic representation of the crossbridge cycle: the myosin cross bridge is bound to actin in rigor 45°- position - "down" (1) ATP binds which leads to very fast dissociation from actin (2)]. The hydrolysis of ATP to ADP and a phosphate ion leads to a return of the myosin cross bridge to the 90° "up" position (3) whereupon it rebinds to actin (4). This leads to release of the products and return to [1]. In the last step actin is "rowed" past myosin.
X-ray fibre diffraction pattern
. The fibre axis is at right angles to the x-ray beam. Note the regular layer lines (with a repeat of (1/430Å) which arise from the helical array of myosin cross bridges. The third order (1/143.5Å) meridional reflexion corresponds to the repeat distance between cross-bridges. (This x-ray fibre diagram was obtained using a rotating anode x-ray generator and mirror-monochromator optics).")

Fig 2: X-ray fibre diffraction pattern of a living frog muscle (Huxley & Brown, 1967). The fibre axis is at right angles to the x-ray beam. Note the regular layer lines (with a repeat of (1/430Å) which arise from the helical array of myosin cross bridges. The third order (1/143.5Å) meridional reflexion corresponds to the repeat distance between cross-bridges. (This x-ray fibre diagram was obtained using a rotating anode x-ray generator and mirror-monochromator optics).
X-ray fibre diffraction pattern of a living frog muscle (Huxley & Brown, 1967). The fibre axis is at right angles to the x-ray beam. Note the regular layer lines (with a repeat of (1/430Å) which arise from the helical array of myosin cross bridges. The third order (1/143.5Å) meridional reflexion corresponds to the repeat distance between cross-bridges. (This x-ray fibre diagram was obtained using a rotating anode x-ray generator and mirror-monochromator optics).
Only the distal part of the cross bridge moves
 indicated that the scheme shown in Fig 1 needed revision: only the distal part of the cross bridge moves (Cooke 1986).")
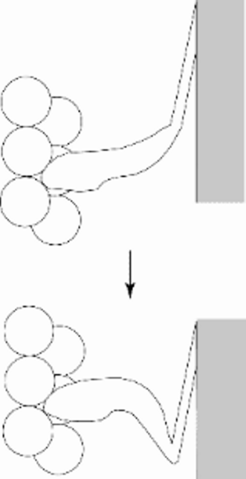
© Kenneth C. Holmes
Fig 6: Numerous experiments (mostly negative) indicated that the scheme shown in Fig 1 needed revision: only the distal part of the cross bridge moves (Cooke 1986).
© Kenneth C. Holmes
Numerous experiments indicated that the scheme shown in Fig 1 needed revision: only the distal part of the cross bridge moves (Cooke, 1986).
The structure of the actin myosin complex
 : shown are (right) five actin molecules in an actin helix (Holmes et al. 1990) and (left) a myosin cross-bridge (S1)(Rayment et al. 1993): shown are: 25K fragment (green); 50K upper fragment (red); 50K lower fragment (white); the disordered chain between the 50K domain and the 20K domain is shown as a yellow loop - note this loop has been modeled; the first part of the 20K domain including the SH2 helix (until 699) is shown light blue; the SH1 helix, converter domain and the C-terminal helix - \"the neck\" (dark blue): the regulatory light chain (magenta) and the essential light chain (yellow). Plates prepared with GRASP (Nicholls et al. 1991). This is the \"end\" state.")

Fig 7a: The structure of the actin myosin complex (Rayment et al. 1993; Schroeder et al. 1993) : shown are (right) five actin molecules in an actin helix (Holmes et al. 1990) and (left) a myosin cross-bridge (S1)(Rayment et al. 1993): shown are: 25K fragment (green); 50K upper fragment (red); 50K lower fragment (white); the disordered chain between the 50K domain and the 20K domain is shown as a yellow loop - note this loop has been modeled; the first part of the 20K domain including the SH2 helix (until 699) is shown light blue; the SH1 helix, converter domain and the C-terminal helix - "the neck" (dark blue): the regulatory light chain (magenta) and the essential light chain (yellow). Plates prepared with GRASP (Nicholls et al. 1991). This is the "end" state.
The structure of the actin myosin complex (Rayment et al. 1993; Schroeder et al. 1993) : shown are (right) five actin molecules in an actin helix (Holmes et al. 1990) and (left) a myosin cross-bridge (S1)(Rayment et al. 1993): shown are: 25K fragment (green); 50K upper fragment (red); 50K lower fragment (white); the disordered chain between the 50K domain and the 20K domain is shown as a yellow loop - note this loop has been modeled; the first part of the 20K domain including the SH2 helix (until 699) is shown light blue; the SH1 helix, converter domain and the C-terminal helix - "the neck" (dark blue): the regulatory light chain (magenta) and the essential light chain (yellow). Plates prepared with GRASP (Nicholls et al. 1991). This is the "end" state.
A reconstruction of the beginning state
. The missing \"neck\" or lever arm has been modeled from chicken S1 data. Note the 70° rotation of the lever arm. The rotation of the lever arm is controlled by the bending out of a helix (shown white) which contacts the g-phosphate at its inner end (the so called\"switch-2\" region). The distal end of the lever arm moves about 12nm between the two states.")

Fig 7b: A reconstruction of the "beginning" state from the crystallographic data on the dictyostelium construct truncated at 761 and complexed with ADP.vanadate (Smith and Rayment 1996). The missing "neck" or lever arm has been modeled from chicken S1 data. Note the 70° rotation of the lever arm. The rotation of the lever arm is controlled by the bending out of a helix (shown white) which contacts the g-phosphate at its inner end (the so called"switch-2" region). The distal end of the lever arm moves about 12nm between the two states.
A reconstruction of the "beginning" state from the crystallographic data on the dictyostelium construct truncated at 761 and complexed with ADP.vanadate (Smith and Rayment 1996). The missing "neck" or lever arm has been modeled from chicken S1 data. Note the 70° rotation of the lever arm. The rotation of the lever arm is controlled by the bending out of a helix (shown white) which contacts the g-phosphate at its inner end (the so called"switch-2" region). The distal end of the lever arm moves about 12nm between the two states.