Verfolgungsjagd in virtuellen Welten – Verarbeitung visueller Bewegung im Gehirn des Zebrabärblings
Forschungsbericht (importiert) 2012 - Max Planck Institut für medizinische Forschung
Lebenswichtiges Bewegungssehen
Wir werden von Bewegung geleitet – oder abgelenkt. Man schaut an einem windstillen Tag in den ruhigen Garten. Plötzlich, irgendwo, regt sich etwas: vielleicht ein in den Ästen kletterndes Eichhörnchen. Nichts zieht unseren aufmerksamen Blick so auf sich wie Bewegung. Im Straßenverkehr dient uns das Bewegungssehen in der Peripherie als Warnsignal. In Sekundenbruchteilen ermittelt unser Gehirn, ob sich ein Objekt auf Kollisionskurs befindet und uns zum Ausweichen zwingt. Und wenn unser Blick einem Objekt in Bewegung folgt, steuert das Sehsystem den Muskelapparat unserer Augen, um das Objekt in den zentralen Bereich der Netzhaut (Retina) zu bringen und dort festzuhalten. Oft geht dieser “visuelle Greifreflex” einer zielgerichteten Bewegung der Hände oder des Körpers voraus, wenn wir den Gegenstand z. B. abfangen wollen, wie der Torwart einen Fußball. Im Tierreich ist dieses visuell gesteuerte Folgeverhalten in jedem Winkel zu beobachten und es erschließt sich uns leicht Ziel und Zweck dieser Jagd. Das Ziel: ein anderes Tier. Der Zweck: Nahrungsaufnahme oder Fortpflanzung. Die lebenswichtige Bedeutung dieser Handlungsmuster lässt erahnen, dass die dem Folgeverhalten zugrunde liegenden neuronalen Schaltkreise effizient und zuverlässig arbeiten müssen, um das Überleben zu ermöglichen.
Die visuelle Verarbeitung von Bewegung wird heute als eine speziell entwickelte Funktion des Sehsystems verstanden, die auf unterschiedlichen Stufen stattfindet und sich in den Aktivitätsmustern von Neuronen manifestiert, welche spezifisch auf bewegte Reize reagieren. Solche bewegungsempfindlichen Nervenzellen findet man bereits in der Retina, aber auch in weiten Teilen der Hirnrinde (Kortex) sowie in tiefer liegenden Sehzentren, wie beispielsweise den oberen Hügelchen (Colliculi Superiores). Diese dem Dach (Tektum) des Mittelhirns entsprechende Struktur erhält direkte Eingänge von Projektionsneuronen der Retina, und zwar dergestalt, dass räumlich benachbarte Reize in unserem Gesichtsfeld auch benachbarte Regionen auf der Oberfläche des Tektums aktivieren (retinotope Abbildung). Zugleich enthalten die tieferen Schichten des Tektums Neuronen, welche Augen- und Kopfbewegungen auslösen können mit dem Ergebnis, dass der Blick auf die der retinotopen Karte entsprechenden Stelle gerichtet wird. Über diese mehr oder weniger direkte Kopplung von Sensorik und Motorik hinaus werden dem Tektum wichtige Funktionen bei der Auswahl und Bewertung von konkurrierenden Zielobjekten und der Aufmerksamkeitssteuerung zugeschrieben. Das bisherige Verständnis dieser Schaltkreise ist einer Vielzahl von Experimenten zu verdanken, in denen zumeist einzelne Zellen mittels feiner Elektroden untersucht wurden. Fundamentale Weiterentwicklungen in der Fluoreszenzmikroskopie ermöglichen es heute, die räumliche und zeitliche Verteilung von Aktivitätsmustern in Hunderten von Nervenzellen an der Oberfläche des Gehirns gleichzeitig zu „filmen“ [1, 2], doch leider ist dieser Ansatz für tiefer liegende Zentren des Bewegungssehens in den meisten Wirbeltieren nicht möglich.
 Larve, 7 Tage alt, Länge etwa 4 mm.
(B) Grün-fluoreszierendes Protein (GFP) markiert die Projektion ausgehend von der Ganglionzellschicht der Retina zum Tektum.
(C) Expression von GFP im Tektum.
(D) Färbung der temporalen und nasalen Retina.
(E) Detailaufnahme des Tektums, mit erkennbaren Schichten bereits in frühen Stadien (7 Tage alte Larve).")
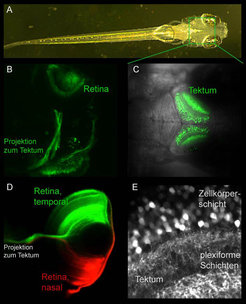
Abb. 1: Sehzentren im Zebrabärbling.
(A) Larve, 7 Tage alt, Länge etwa 4 mm.
(B) Grün-fluoreszierendes Protein (GFP) markiert die Projektion ausgehend von der Ganglionzellschicht der Retina zum Tektum.
(C) Expression von GFP im Tektum.
(D) Färbung der temporalen und nasalen Retina.
(E) Detailaufnahme des Tektums, mit erkennbaren Schichten bereits in frühen Stadien (7 Tage alte Larve).
Ein „gläserner“ Jäger gewährt Einblick in sein Gehirn – Zebrabärbling als Modellsystem
Die Larve des Zebrabärblings, salopp auch Zebrafisch genannt, ist eine glückliche Ausnahme. Die Larve und ihr Gehirn sind klein; im Alter von wenigen Tagen misst die Larve etwa vier Millimeter und ihr Gehirn ist an der dicksten Stelle nicht tiefer als 0,5 mm (Abb. 1). Die Haut der Zebrafischlarve ist durchsichtig, sodass praktisch alle Bereiche ihres Gehirns unter Verwendung der Mehrphotonen-Fluoreszenzmikroskopie mit zellulärer Auflösung „durchleuchtet“ werden können. Um außerdem noch das Feuern von Neuronen sichtbar zu machen, können Wissenschaftler auf ein großes Spektrum molekularbiologischer Methoden zurückgreifen. Damit können Nervenzellen gezielt zur Anreicherung von fluoreszierenden, Kalzium-bindenden Proteinen veranlasst werden, die die mit neuronaler Aktivität einhergehenden Kalziumsignale sichtbar machen [3]. Interessant wird die Zebrafischlarve für die Neurobiologie vor allem, weil sie schon in frühen Stadien über Sinnessysteme verfügt, die denen anderer Wirbeltiere prinzipiell gleichen. So sind z. B. die Augen und die Sehzentren des Mittelhirns der Larve den subkortikalen Sehzentren der Säuger im Aufbau ähnlich (Abb. 1). Die Summe dieser Vorteile gibt Anlass zur Hoffnung, dass in der Zebrafischlarve erreicht werden kann, was ein Langzeitziel der Hirnforschung ist, nämlich das Zusammenwirken aller Neurone im intakten Gehirn zu verstehen.
Phantomjagd: Die virtuelle Welt der Zebrafischlarve
Das gut entwickelte Sehsystem macht die Zebrafischlarve zu einem beeindruckenden Jäger, der kleine Wimpertierchen zielstrebig verfolgt und fängt [4]. Die Aufgabe ist nicht einfach, da das Beutetier kaum kleiner als das Maul der Larve ist und häufig seine Richtung wechselt. Entfernt erinnert das Problem an den Versuch, einen Schmetterling mit einem zu kleinen Netz zu fangen: nur ein schnelles Reaktionsvermögen und häufige Richtungsanpassungen des Fängers führen zum Erfolg.
 Einzelaufnahmen der Larve beim Beutefang. Eine Fangsequenz dauert wenige Sekunden.
(B) Quantifizierung der Augen- und Schwanzbewegungen.
(C) Kinematische Studien der Drehbewegung (links) und der Vorwärtsbewegung (rechts) der Larve beim Beutefangverhalten.
(D) Zeitverlauf der Ausrichtung der Larve während einer Beutefangsequenz. Darunter: Zeitverlauf der entsprechenden Schwanzbewegungen, mit denen die Larve ihre Ausrichtung steuert.
(E) Virtuelles Beutefangverhalten: Die teilweise eingebettete Larve betrachtet virtuelle Beute auf einer kleinen Leinwand. Sobald sie gerichtet schwimmt, wird eine Drehbewegung durch das Verschieben der projizierten Umgebung simuliert.
(F) Die virtuelle Beute aus der Sicht der Larve.")
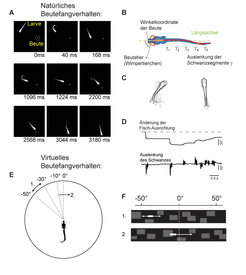
Abb. 2: Beutefangverhalten der Zebrabärblingslarve
(A) Einzelaufnahmen der Larve beim Beutefang. Eine Fangsequenz dauert wenige Sekunden.
(B) Quantifizierung der Augen- und Schwanzbewegungen.
(C) Kinematische Studien der Drehbewegung (links) und der Vorwärtsbewegung (rechts) der Larve beim Beutefangverhalten.
(D) Zeitverlauf der Ausrichtung der Larve während einer Beutefangsequenz. Darunter: Zeitverlauf der entsprechenden Schwanzbewegungen, mit denen die Larve ihre Ausrichtung steuert.
(E) Virtuelles Beutefangverhalten: Die teilweise eingebettete Larve betrachtet virtuelle Beute auf einer kleinen Leinwand. Sobald sie gerichtet schwimmt, wird eine Drehbewegung durch das Verschieben der projizierten Umgebung simuliert.
(F) Die virtuelle Beute aus der Sicht der Larve.
Um die zielgerichteten Bewegungsmuster der Larve quantitativ zu erfassen, untersucht die Forschungsgruppe um Johann Bollmann am Max-Planck-Institut für medizinische Forschung das Schwimmverhalten mit Hilfe von Hochgeschwindigkeitskameras, wie sie auch bei Crashtests im Fahrzeugbau zum Einsatz kommen (Abb. 2). Anschließend verwenden die Forscher die daraus gewonnenen Informationen, um eine virtuelle Welt für den Zebrafisch zu errichten, in der die Beute nur noch aus projizierten Bildern besteht. Ist die Larve in dieser Welt mittels etwas Agarose eingebettet, so führt sie doch Schwimmbewegungen in Richtung des virtuellen Wimpertierchens aus. Werden auch noch die Schwimmversuche in Echtzeit optisch gemessen und in entsprechende Änderungen der virtuellen Umgebung umgesetzt, so führt die Larve ganze Sequenzen ihres natürlichen Beutefangverhaltens aus. Was zunächst wie ein Spiel klingt, bietet tatsächlich immense Vorteile beim Studium des visuellen Systems. So kann zum Beispiel durch exakt gesteuerte Verzögerungen oder Verschiebungen der virtuellen Beute getestet werden, wie das Sehsystem sehr schnell auf unerwartete Fehler bei der Bewegungsausführung reagiert. Und da die Larve in der virtuellen Beutefangwelt ganze Schwimmabfolgen ausführt, ohne sich tatsächlich fortzubewegen, eröffnet diese Technik die Möglichkeit, den zugrunde liegenden Informationsfluss der elektrischen Signale im Nervensystem unter dem Fluoreszenzmikroskop messbar zu machen.
Arbeitsteilung im Gehirn: Neuronen, die Bewegung „sehen“
 Expression von GFP (grün) in einem spezifischen Zelltyp des Tektums hebt die Schichtung dieses Sehzentrums hervor.
(B) Derselbe Zelltyp wie in (A), aber hier mit einem Kalzium-empfindlichen Protein gefärbt.
(C) Kalziumsignale gemessen im Zellkörper zeigen Richtungsempfindlichkeit: das rot umrandete Neuron in B bevorzugt Bewegung nach anterior (nach vorn gerichtet).
(D) Zwei strukturell unterschiedliche Neuronen (Zelltyp 1 und 2; jeweils rot gefärbt) weisen unterschiedliche Vorzugsrichtungen auf. Spuren über den Aufnahmen stellen die Feuerraten in den abgebildeten Neuronen bei unterschiedlichen Bewegungsrichtungen dar.")
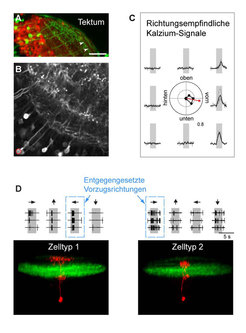
Abb. 3: Richtungsempfindliche Zelltypen im Tektum
(A) Expression von GFP (grün) in einem spezifischen Zelltyp des Tektums hebt die Schichtung dieses Sehzentrums hervor.
(B) Derselbe Zelltyp wie in (A), aber hier mit einem Kalzium-empfindlichen Protein gefärbt.
(C) Kalziumsignale gemessen im Zellkörper zeigen Richtungsempfindlichkeit: das rot umrandete Neuron in B bevorzugt Bewegung nach anterior (nach vorn gerichtet).
(D) Zwei strukturell unterschiedliche Neuronen (Zelltyp 1 und 2; jeweils rot gefärbt) weisen unterschiedliche Vorzugsrichtungen auf. Spuren über den Aufnahmen stellen die Feuerraten in den abgebildeten Neuronen bei unterschiedlichen Bewegungsrichtungen dar.
Ein wichtiges Ziel zum Verständnis des Gehirns ist es zu verstehen, welche Aufgaben verschiedene Nervenzellgruppen, sogenannte Schaltkreismotive, aufgrund ihres synaptischen Verknüpfungsmusters übernehmen könnten. So wird z. B. seit langem die Frage diskutiert, welche neuronalen Mechanismen dafür verantwortlich sind, dass die Bewegungsrichtung eines Reizes in Form unterschiedlicher neuronaler Feuerraten verschlüsselt wird, was die „Richtungsempfindlichkeit“ eines Neurons bezeichnet.
Im Gehirn des Zebrafisches haben die Heidelberger Max-Planck-Forscher nun ein neues Schaltkreismotiv entdeckt, das mit der Verarbeitung von visuellen Bewegungsrichtungen befasst ist [3]. Wie auch in vielen anderen Hirn-Arealen findet man im Tektum richtungsempfindliche Nervenzellen, ohne ihre Verschaltung zu kennen. Gemeinsam mit der Gruppe von Soojin Ryu im selben Institut schleusten die Wissenschaftler ein Gen für den molekularen Kalziumsensor „GCaMP3“ so in die Zebrafischlarve ein, dass es nur in einer geringen Zahl von Nervenzellen angeschaltet wird und diese zum Leuchten bringt. Mit Hilfe der Mehrphotonenmikroskopie ließ sich damit die Richtungsempfindlichkeit einzelner Zellen bestimmen (Abb. 3).
Unter Verwendung feinster Glaspipetten konnten identifizierte Zellen anschließend mit einem roten Fluoreszenzfarbstoff gefüllt werden, wodurch innerhalb weniger Minuten ein detailliertes Bild der betreffenden Zelle und ihres dendritischen Verzweigungsmusters sichtbar wurde. So ließen sich zwei strukturell verschiedene, richtungsempfindliche Zelltypen mit entgegengesetzten Vorzugsrichtungen nachweisen. Überraschenderweise liegen die dendritischen Verzweigungen dieser zwei Zelltypen geschichtet in verschiedenen Lagen des Tektums.
 Ein 3-dimensionales Volumen aus dem Rückenmark der Larve, aufgenommen durch serielle Rasterelektronenmikroskopie (SBEM). Ein einzelner Bildpunkt entspricht 10 x 10 x 23 Nanometern.
(B) Diese Auflösung reicht aus, um die Kontaktstellen (Synapsen) zwischen Nervenzellen zu erkennen. Links ist ein Kontakt zwischen einem Axon und einem Zellkörper, rechts ein Kontakt zwischen zwei Axonen zu sehen.")
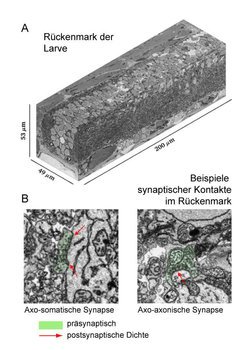
Abb. 4: Serielle blockface-Rasterelektronenmikroskopie im Rückenmark
(A) Ein 3-dimensionales Volumen aus dem Rückenmark der Larve, aufgenommen durch serielle Rasterelektronenmikroskopie (SBEM). Ein einzelner Bildpunkt entspricht 10 x 10 x 23 Nanometern.
(B) Diese Auflösung reicht aus, um die Kontaktstellen (Synapsen) zwischen Nervenzellen zu erkennen. Links ist ein Kontakt zwischen einem Axon und einem Zellkörper, rechts ein Kontakt zwischen zwei Axonen zu sehen.
In weiterführenden Untersuchungen konnten die Heidelberger Forscher nachweisen, dass richtungsempfindliche Signale von Projektionsneuronen der Retina ebenfalls in diesen Schichten eintreffen, sodass hier erstmals der Nachweis für eine schichtspezifische Verarbeitung von Richtungsempfindlichkeit in einem Sehzentrum jenseits des Auges gelang [3]. Für Erkenntnisse der Art, wie die räumliche Struktur eines Schaltkreises seine Funktion bedingt, ist die Zebrafischlarve aufgrund ihrer Kompaktheit, der genetischen Zugänglichkeit und ausgereiften Visuomotorik ein hervorragendes Untersuchungsmodell [5, 6].
Mehr als eine Handvoll Zellen: Die vollständige Rekonstruktion des Zebrafisch-Gehirns
Die Auflösung der Lichtmikroskopie reicht auch in diesem Tiermodell nicht aus, um die feinen Verästelungen und synaptischen Kontakte aller Neuronen in Gänze sichtbar zu machen. Das Ziel, einen vollständigen Verschaltungsplan des Gehirns dieses kleinen Jägers zu erhalten, erfordert das Auflösungsvermögen des Elektronenmikroskops (Abb. 4). Zusammen mit Winfried Denk arbeiten die Forscher am Heidelberger Max-Planck-Institut daran, mit Hilfe der seriellen blockface-Rasterelektronenmikroskopie (SBEM) [7] das Gehirn der Zebrafischlarve Schnitt für Schnitt mit höchster Auflösung abzufotografieren, um anschließend die Drähte und Verschaltungen des Gehirns am Computer zu rekonstruieren [8]. Solch ein umfassender Verschaltungsplan hätte unschätzbaren Wert: Wir könnten besser verstehen, welche Wege der Informationsfluss in biologischen Netzwerken nimmt, wenn das Gehirn unser Verhalten steuert.
Literaturhinweise
Nature Reviews Neuroscience 13, 351-358 (2012)