Introduction to Muscle Contraction, Part 1
What makes muscles shorten?
The contraction of voluntary muscles in all animals takes place by the mutual sliding of two sets of interdigitating filaments: thick (containing the protein myosin) and thin (containing the protein actin) organized in sarcomeres each a few microns long which give muscle its cross striated appearance in the microscope (Fig 1). The relative sliding of thick and thin filaments is brought about by cross bridges, parts of the myosin molecules which sticks out from the myosin filaments and interact cyclically with the thin filaments, transporting them by a kind of rowing action. During the process ATP (adenosine triphosphate) is hydrolyzed to ADP (adenosine diphosphate), the hydrolysis of ATP provides the energy.
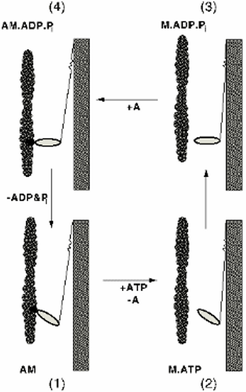
The protein myosin was discovered more than 100 years ago by Kühne (Kühne 1864). ATP was discovered by Lohmann in our Institute (Lohmann 1931) and Lyubimova and Engelhardt (Engelhardt and Ljubimowa 1939) showed that the ATP was hydrolyzed by myosin and is the immediate energy source for muscle. Working with Albert Szent Gyorgyi in Szeged, Straub discovered that myosin was actually two proteins, myosin and actin (Straub 1943). ATP was also shown to be a relaxing factor - i.e. ATP also dissociates actin and myosin. Moreover, Albert Szent Gyorgyi was able to show that glygerol-treated muscle fibres, containing only actin and myosin shorten on adding ATP. A similar result was obtained by HH Weber using artificial threads of actomyosin. The dichotomy of the action of ATP (a relaxing factor that drives contraction) remained an enigma to be explained later by Lymn and Taylor. In the mean time, the myosin molecule was characterized and was shown to consist of two heavy chains and two light chains. A soluble proteolytic fragment of myosin, heavy mero-myosin (which contains the globular heads of myosin) contains the ATP-ase activity (Szent-Gyorgyi 1953), the rest of the molecule forming a long a-helical coiled-coil. The ATPase activity was later shown to reside in the head itself (Margossian and Lowey 1973) - often called S1 - which constitutes the morphological "cross-bridge".

© Kenneth C. Holmes
Fig 2
© Kenneth C. Holmes
The first molecular theories, which appeared in the 30s, were based on polymer science. They proposed that there was a rubber-like shortening of myosin filaments brought about by altering the state of ionization of the myosin. This aberration was corrected by the seminal works of HE Huxley (Huxley and Hanson 1954) and AF Huxley (Huxley and Niedergerke 1954) which showed that sarcomeres contained two sets of filaments (thick and thin) which glided over each other without altering their length. Hasselbach showed that the thick filaments contain the protein myosin. The question naturally arose; what made the filaments glide? Projections from the thick filaments, the myosin cross bridges, were discovered by electron microscopy (Huxley 1957; Huxley 1958) and subsequently shown both to be the site of the ATPase and also to be the motor elements producing force and movement between the filaments. Two conformations of the cross-bridge could be detected in insect flight muscle (Reedy et al. 1965). Progress was then rapid so that at a historic Cold Spring Harbor Symposium in 1972 the outline of the molecular mechanism of muscle contraction could be enunciated. The cross bridge was thought to bind to actin in an initial (90°) conformation, go over to a angled (45°) conformation and then release (Huxley 1969) (Lymn and Taylor 1971). For each cycle of activity one ATP would be hydrolyzed. The actual movement could be measured by physiological experiments on contracting muscle and was shown to be about 80-100Å (Huxley and Simmons 1971). Since the cross-bridge was known to be an elongated structure, such a distance could be accommodated by a rotating or swinging cross-bridge model.