Introduction to Muscle Contraction, Part 1
A primary aim of the Department of Biophysics since its foundation in 1968 has been to understand in physiochemical terms the molecular mechanism by which muscles produce force. Muscle contraction involves the cyclic interaction of the proteins myosin and actin, often pictured as the rowing of the myosin and actin filaments past each other, using the hydrolysis of ATP as a source of energy. We study the structures of actin and myosin at atomic and near-atomic resolution by protein crystallography, electron microscopy and X-ray fibre diffraction.
As a supplementary technique for studying mobility we also use NMR. We study myosins from various sources and with a varity of bound nucleotide analogues, also in combination with site-directed mutagenesis. A biochemistry group which specializes in the expression of proteins in the cellular slime mould dictyostelium uses enzyme kinetics and in vitro motility assays to guage the effects of mutagenesis. The most interesting cases are then analysed by x-ray crystallography. This project is part of an international collaboration.
The last two years have seen dramatic progress in our understanding of the molecular basis of muscle contraction and indeed we now have considerable understanding of the processes involved in myosin-based motility. The crystallography goup studies other proteins as well, in particular on those which, like myosin, involve the processing of nucleotides.
Research on muscle contraction goes back to the Greeks
Understanding muscle contraction answers one of the fundamental questions posed in classical times, namely the nature of the spiritus animalis. The spiritus animalis was an intrinsic property of living things. Erasistratus (3rd century B.C.) of the Alexandrian school associated the spiritus animalis with the muscles. The pneuma was thought to course along the nerves and make the muscles swell and shorten. In the beginning of the 2nd. century A.D. Galen, the last classical physiologist took over and expanded these ideas introducing a primitive metabolism involving the four humours. Furthermore, Galen made a detailed anatomical examination of muscles and understood that they worked in antagonistic pairs, and that the heart was a muscle for pushing blood into the arteries. In the ensuing millenium nothing much happened and even Galen's insight that muscles pull rather than push seems to have been forgotten, since at the beginning of the ¢16, on the basis of his own anatomical examinations Leonardo da Vinci wrote:
Perchè l'ufizio del musculo è di tirare e non di spingere.
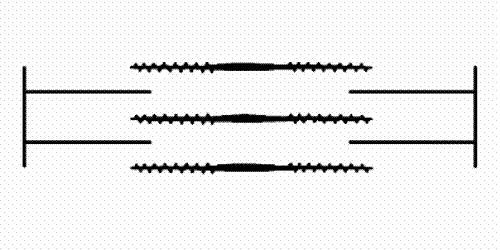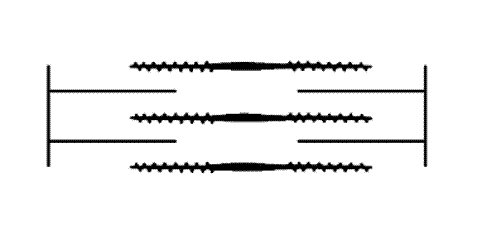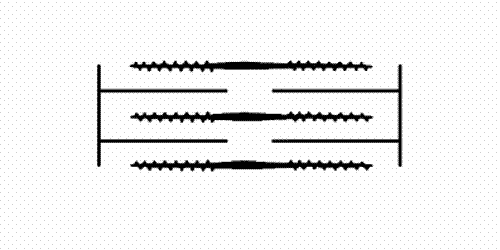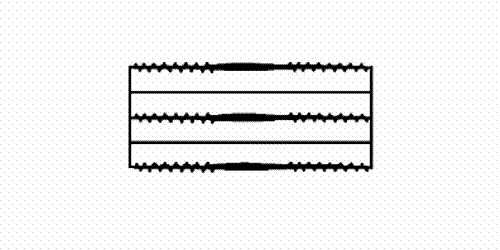
 filaments and actin (thin) filaments. As muscle contracts the sets of filaments move past each other. The movement is produced by the myosin cross-bridges which interact cyclically with the actin filaments and move them by a kind of rowing action. A muscle fibril consists of many thousand sacomeres arranged in series.")
© Kenneth C. Holmes
Figure 1: The sarcomere consists of interdigitating myosin (thick) filaments and actin (thin) filaments. As muscle contracts the sets of filaments move past each other. The movement is produced by the myosin cross-bridges which interact cyclically with the actin filaments and move them by a kind of rowing action. A muscle fibril consists of many thousand sacomeres arranged in series.
© Kenneth C. Holmes
A few years later Vesalius used the phrase Machina Carnis to underline the fact that the production of force resided in the flesh (muscle) itself, and in the early ¢16 Descartes proposed a neuromuscular machine not unlike that of Erasistratus: the nerves carry a fluid from the pineal gland (the seat of the soul) to the muscles which makes them swell and shorten. A little later, Swammerdam was to show that muscles contract at constant volume which invalidated this whole class of pneumatic theories. However, other mechanical models were soon proposed. Alongside such mechanical thinking, however, vitalism survived into the ¢19th., and one needed the whole fabric of metabolic biochemistry and thermodynamics to support the concept that muscle is a chemical machine driven by isothermal combustion which was first articulated by von Helmholtz.
What makes muscles shorten?
The contraction of voluntary muscles in all animals takes place by the mutual sliding of two sets of interdigitating filaments: thick (containing the protein myosin) and thin (containing the protein actin) organized in sarcomeres each a few microns long which give muscle its cross striated appearance in the microscope (Fig 1). The relative sliding of thick and thin filaments is brought about by cross bridges, parts of the myosin molecules which sticks out from the myosin filaments and interact cyclically with the thin filaments, transporting them by a kind of rowing action. During the process ATP (adenosine triphosphate) is hydrolyzed to ADP (adenosine diphosphate), the hydrolysis of ATP provides the energy.
The protein myosin was discovered more than 100 years ago by Kühne (Kühne 1864). ATP was discovered by Lohmann in our Institute (Lohmann 1931) and Lyubimova and Engelhardt (Engelhardt and Ljubimowa 1939) showed that the ATP was hydrolyzed by myosin and is the immediate energy source for muscle. Working with Albert Szent Gyorgyi in Szeged, Straub discovered that myosin was actually two proteins, myosin and actin (Straub 1943). ATP was also shown to be a relaxing factor - i.e. ATP also dissociates actin and myosin. Moreover, Albert Szent Gyorgyi was able to show that glygerol-treated muscle fibres, containing only actin and myosin shorten on adding ATP. A similar result was obtained by HH Weber using artificial threads of actomyosin. The dichotomy of the action of ATP (a relaxing factor that drives contraction) remained an enigma to be explained later by Lymn and Taylor. In the mean time, the myosin molecule was characterized and was shown to consist of two heavy chains and two light chains. A soluble proteolytic fragment of myosin, heavy mero-myosin (which contains the globular heads of myosin) contains the ATP-ase activity (Szent-Gyorgyi 1953), the rest of the molecule forming a long a-helical coiled-coil. The ATPase activity was later shown to reside in the head itself (Margossian and Lowey 1973) - often called S1 - which constitutes the morphological "cross-bridge".

© Kenneth C. Holmes
Fig 2
© Kenneth C. Holmes
The first molecular theories, which appeared in the 30s, were based on polymer science. They proposed that there was a rubber-like shortening of myosin filaments brought about by altering the state of ionization of the myosin. This aberration was corrected by the seminal works of HE Huxley (Huxley and Hanson 1954) and AF Huxley (Huxley and Niedergerke 1954) which showed that sarcomeres contained two sets of filaments (thick and thin) which glided over each other without altering their length. Hasselbach showed that the thick filaments contain the protein myosin. The question naturally arose; what made the filaments glide? Projections from the thick filaments, the myosin cross bridges, were discovered by electron microscopy (Huxley 1957; Huxley 1958) and subsequently shown both to be the site of the ATPase and also to be the motor elements producing force and movement between the filaments. Two conformations of the cross-bridge could be detected in insect flight muscle (Reedy et al. 1965). Progress was then rapid so that at a historic Cold Spring Harbor Symposium in 1972 the outline of the molecular mechanism of muscle contraction could be enunciated. The cross bridge was thought to bind to actin in an initial (90°) conformation, go over to a angled (45°) conformation and then release (Huxley 1969) (Lymn and Taylor 1971). For each cycle of activity one ATP would be hydrolyzed. The actual movement could be measured by physiological experiments on contracting muscle and was shown to be about 80-100Å (Huxley and Simmons 1971). Since the cross-bridge was known to be an elongated structure, such a distance could be accommodated by a rotating or swinging cross-bridge model.
Why did it take so long to work out?
To understand the chemical events which drive muscle one needs to know the protein structures involved at atomic resolution. Muscle is made of massive arrays of macromolecules. How does one get data from such systems? A great deal of technology has been invested in this problem, which has also driven technology. Some early insight was provided by light microscopy. However, the first radical new insight came from electron microscopy. More recently, the structures of the component molecules have been determined x-ray crystallography at atomic resolution. These results now allow us to describe in some detail how the hydrolysis of ATP by the component proteins actin and myosin leads to movement.
An understanding of muscle contraction is an important example of the success of protein crystallography, in particular when used in conjunction with high resolution electron microscopy.
Time-resolved X-ray Diffraction from Frog Muscle
Excised living frog muscles will continue to contract for may hours if bathed in Ringers solution and stimulated electrically. HE Huxley showed that living frog muscles give a detailed low-angle x-ray fibre diagrams with a series of layer-lines arising from the helical arrangement of cross-bridges. One of these has a strong meridional peak with a Bragg spacing of 143.5Å arising from the distance between cross bridges along the myosin filament helix. If the cross bridges tilt during contraction, then this reflexion should get weaker. Muscles contract fast, so millisecond time resolution is necessary. Initial attempts to observe this effect with conventional x-ray sources were unable to get enough signal in the short times available. Therefore, the first beam lines for synchrotron radiation as an x-ray source were set up at DESY Hamburg (Rosenbaum et al. 1971). Synchrotron radiation provided the requisite power.
The key experiments were actually carried out in 1980 (Huxley et al. 1981). These finally showed the anticipated changes in intensity of the meridional reflexions. If a contracting muscle is released the intensity of the 14.3.5Å meridional reflexion drops within a few ms to a fraction of its initial value. If the muscle is extended quickly, the intensity is recovered. If one waits at the new length the intensity recovers. These experiments have recently been repeated with very high time resolution using sophisticated mechanics and the excellent two dimensional detectors at Daresbury (Irving et al. 1992) These observations are fully consistent with the swinging cross bridge hypothesis and represent the most important time resolved experiments supporting this class of hypothesis.
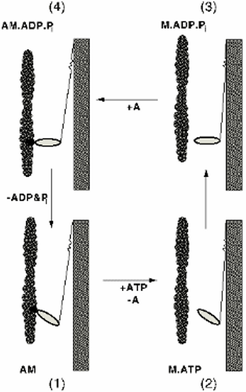
The Cross-Bridge Cycle
In the absence of ATP, the myosin cross-bridge binds tightly to actin filaments. However, it also binds and hydrolyses ATP. ATP binding brings about a rapid dissociation of the cross bridge from actin. Thus the cross bridge can bind either actin or ATP but to both only transiently. The presence of the ATP γ-phosphate is crucial for dissociation since ADP alone has little effect. Solution kinetic observations were very important in establishing the relationship between the hydrolysis of ATP and the generation of force. A key feature of this process is the observation that transduction of the chemical energy released by the hydrolysis of ATP into directed mechanical force should occur during product release (ADP and inorganic phosphate, Π) rather than during the hydrolysis step itself (Lymn and Taylor 1971). Without actin, myosin is product-inhibited and is a poor ATPase.
Mg-ATP rapidly dissociates the actomyosin complex on binding to the ATPase site of myosin; myosin then hydrolyzes ATP and forms a stable myosin-products complex; actin recombines with this complex and dissociates the products, initially the γ-phosphate ion thereby forming the original actin-myosin complex. Force is generated during the last step.

Kenneth C. Holmes
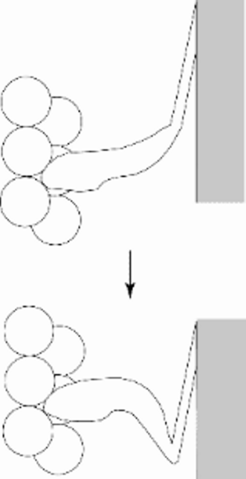
Figure 3: Numerous experiments (mostly negative) indicated that the scheme shown in Fig 1 needed revision: only the distal part of the cross bridge moves (Cooke 1986).
Kenneth C. Holmes
Although the swinging myosin cross bridge hypothesis of muscle contraction had become the textbook norm by 1972 it has proved remarkably difficult to catch a bridge in flagranti delicto (see review by Cooke, 1986). In fact the hypothesis was never very clear about how the cross-bridge moved on actin and had been modified over the years into a swinging lever arm hypothesis in which the bulk of the cross bridge is envisaged to bind to actin with a more or less fixed geometry and only the distal (C-terminal) part of the myosin molecule moves (Holmes 1997). A swinging lever arm explains why substantial changes in the cross bridge orientation were not visible: only a small fraction of the cross-bridge mass moves. Furthermore, it gradually became clear that the proportion of cross bridges taking part in a contraction at any one time was only a small fraction of the total, making the registration of active cross bridge movement doubly difficult.
Atomic Structures of Actin and Myosin
Actin (thin filament) fibres are helical polymers of g-actin (globular-actin) . The structure of monomeric actin which contains 365 residues and has a molecular weight of 42,000 was solved by protein crystallography as a 1:1 complex with the enzyme DNase I (Kabsch et al. 1990). Orientated gels of actin fibres (f-actin), a helical copolymer of actin which has 13 molecules in 6 turns repeating every 360Å, yield x-ray fibre diagrams to about 6Å resolution. It was possible to determine the orientation of the g-actin monomer which best accounted for the f-actin fibre diagram (Holmes et al. 1990) and thus arrive at an atomic model of the actin filament (Lorenz et al. 1993).
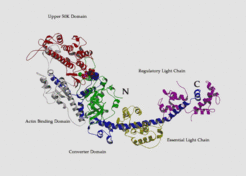
. The structure of chicken S1 has been solved by x-ray crystallography (Rayment et al. 1993) - in the following references to residues in the chicken structure will be prefaced with \"gg\". This study shows the S1 (which has 884 residues) to be tadpole-like in form, with an elongated head, containing a 7-stranded β sheet and numerous associated a-helices forming a deep cleft, with the actin binding sites and nucleotide binding sites on opposite sides of the sheet. The cleft separates two parts of the molecule which are referred to as the 50K upper and 50K lower domains or actin binding domain.")
© Kenneth C. Holmes
The cross-bridges comprise a part of the myosin molecule, namely subfragment-1 of heavy meromyosin (S1). The structure of chicken S1 has been solved by x-ray crystallography (Rayment et al. 1993) - in the following references to residues in the chicken structure will be prefaced with "gg". This study shows the S1 (which has 884 residues) to be tadpole-like in form, with an elongated head, containing a 7-stranded β sheet and numerous associated a-helices forming a deep cleft, with the actin binding sites and nucleotide binding sites on opposite sides of the sheet. The cleft separates two parts of the molecule which are referred to as the 50K upper and 50K lower domains or actin binding domain.
© Kenneth C. Holmes
The cross-bridges comprise a part of the myosin molecule, namely subfragment-1 of heavy meromyosin (S1). The structure of chicken S1 has been solved by x-ray crystallography (Rayment et al. 1993) - in the following references to residues in the chicken structure will be prefaced with "gg". This study shows the S1 (which has 884 residues) to be tadpole-like in form, with an elongated head, containing a 7-stranded β sheet and numerous associated α-helices forming a deep cleft, with the actin binding sites and nucleotide binding sites on opposite sides of the sheet. The cleft separates two parts of the molecule which are referred to as the 50K upper and 50K lower domains or actin binding domain.
The C-terminal tail, sometimes called the neck, which also provides the connection to the thick filament forms an extended α-helix which binds two light chains. The ATP binding site contained the typical P-loop motive which is also found in the G-proteins.
")
© Kenneth C. Holmes
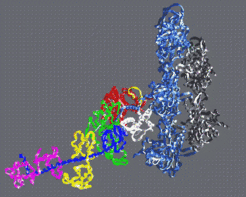
Fig 4: By fitting the atomic structures of f-actin and S1 into three dimensional cryo-electron microscope reconstructions one arrives at an atomic model of the actin myosin complex (Rayment et al. 1993).
© Kenneth C. Holmes
By fitting the atomic structures of f-actin and S1 into three dimensional cryo-electron microscope reconstructions one arrives at an atomic model of the actin myosin complex (Rayment et al. 1993, Fig 4). In particular, this model establishes the spatial orientation of the S1 myosin fragment in the active complex. For example one finds that the cleft in myosin extends from the ATP binding site to the actin binding site and that the opening and closing of this cleft is very likely to provide the communication between the ATP site and the actin binding site. The actin binding site spans the 50K upper and lower domains and the ATP binding site extends from the 50K upper domain into the 50K lower domain. Furthermore, the very extended C-terminal α-helical neck of S1 is ideally placed to be a lever arm. The lever arms joins onto the bulk of the molecule via a small compact converter domain (Houdusse and Cohen 1996) which lies just distal to a broken α-helix containing two reactive thiol groups known as SH1 and SH2. Numerous experiments point to the putative hinge for the lever arm being in the SH1-SH2 region of the molecule (see Holmes 1997 for review).